§ 1. Стадии развития дрожжей и механизм синтеза биомассы
Стадии развития дрожжей. Дрожжевой клетке, как и всем микроорганизмам, присущи четыре основные фазы роста: лагфаза, фаза логарифмического роста, стационарная фаза и фаза отмирания. Однако процесс выращивания дрожжей предусматривает только первые три фазы. Фазы отмирания клеток не должно быть в дрожжевом производстве.
Лагфаза - это период, во время которого внесенные в питательную среду дрожжевые клетки адаптируются к окружающей среде. Их ферментные системы перестраиваются на синтез биомассы. Происходит активный синтез аминокислот, полифосфатов, рибонуклеиновых кислот (РНК). Клетки подготавливаются к почкованию, масса их увеличивается в результате увеличения размеров клеток. Дрожжи в это время не размножаются. Продолжительность этого периода зависит от активности ферментных систем клетки, количества засевного материала, количества и состава питательного субстрата, в который помещены клетки, а также от температуры, рН, аэрации культуральной среды и др. В этот период клетки обладают повышенной чувствительностью к внешним воздействиям.
Фаза логарифмического роста характеризуется высокой активностью размножения клеток. При обеспечении их необходимыми питательными и ростовыми веществами, а также при отсутствии лимита по растворенному кислороду в культуральной среде количество почкующихся клеток быстро увеличивается, достигая 70-80%. Биомасса дрожжей при этом постепенно повышается вследствие образования новых дочерних клеток, которые, вырастая до материнской клетки, начинают почковаться, образуя новую генерацию. После отпочковывания молодой клетки материнские клетки вновь начинают почковаться.
По данным А. Кука, одна дрожжевая клетка способна воспроизвести в среднем 25 новых клеток, а иногда и 40.
В этой фазе наблюдается максимальная скорость роста дрожжевых клеток. Продолжительность периода зависит от количества питательных веществ в среде, аэрации, количества образовавшихся продуктов метаболизма клеток. При периодическом процессе скорость роста постепенно понижается вследствие недостатка источников питания, накопления ингибирующих продуктов или некоторых изменений физических свойств среды. При условии постоянного пополнения среды питательными веществами и выведении из культуральной среды вредных продуктов обмена клеток период логарифмического роста может продолжаться практически бесконечно. На этой особенности микроорганизмов основано непрерывное культивирование микроорганизмов, в частности хлебопекарных дрожжей.
Устойчивость к неблагоприятным условиям внешней среды в этот период повышается.
В стационарной фазе образование новых клеток практически прекращается, заканчивается также и их почкование, так как питательные вещества в дрожжерастильный аппарат не поступают. Для поддержания жизнедеятельности клетки используют оставшиеся в культуральной среде питательные вещества. Клетки увеличиваются в размере, масса их также возрастает. Прирост биомассы в этой фазе может составить 5-10% от массы дрожжей, накопленных в логарифмической фазе. Ферментные системы клеток перестраиваются с активного синтеза биомассы на процессы обмена, поддерживающие их жизнедеятельность. Устойчивость клеток к окружающей среде значительно повышается. Эта фаза соответствует дозреванию клеток в дрожжевом производстве.
Фаза отмирания характеризуется отсутствием роста и размножения микроорганизмов. Масса клеток уменьшается, так как все питательные вещества культуральной среды использованы в предыдущей фазе. Для поддержания жизнедеятельности клетки начинают использовать собственные запасы. Наблюдается автолиз клеток. Качество дрожжей резко ухудшается. Технологическим процессом получения хлебопекарных дрожжей эта фаза не предусматривается, так как дрожжи выделяют из культуральной среды. Она может быть только при аварии, например при поломке сепараторов, когда дозревшие дрожжи длительное время простаивают в дрожжерастильных аппаратах без притока питательных веществ и др.
Для повышения съема биомассы с дрожжерастильного аппарата необходимо технологический процесс вести так, чтобы сократить до минимума период лагфазы и увеличить фазу логарифмического роста клеток. От этих двух фаз зависят в основном выработка и выход дрожжей. Стационарная фаза также влияет на величину этих двух показателей. Однако от правильности ведения процесса в этой фазе в основном зависит качество готового продукта (хлебопекарное достоинство и сохранность дрожжей).
Механизм синтеза биомассы. В основе жизнедеятельности любого организма лежат процессы обмена веществ, для протекания которых требуется постоянный и непрерывный приток энергии.
Большинство организмов для поддержания своей жизни используют энергию, освобождающуюся во время диссимиляции (разрушения) органических веществ, в. первую очередь сахара.
В дрожжевой клетке диссимиляция сахара может происходить либо аэробным путем, т. е. благодаря процессу дыхания (в присутствии кислорода), либо анаэробно, т. е. путем брожения (при отсутствии кислорода или недостаточном количестве его в культуральной среде).
На основании последних достижений в области биологии выявилась предположительная схема синтеза белковых веществ в дрожжевых клетках. Сначала происходит расщепление сахара до пировиноградной кислоты по схеме Эмбдена - Мейергофа - Парнаса (ЭМП), а затем в условиях интенсивной аэрации - по циклу трикарбоновых и дикарбоновых кислот, называемому циклом Кребса.
Конечным продуктом цикла Кребса является щавелевоуксусная кислота, которая, соединяясь с аммиаком, образует аспарагиновую кислоту, являющуюся основой белковой структуры клетки. Под действием фермента пируваткарбоксилазы щавелевоуксусная кислота может образовать вновь пировиноградную кислоту. Эта реакция обратима.
Кроме основного пути распада углеводов через пировиноградную кислоту, лежащую в основе дыхания и брожения, существуют и другие пути окислительного превращения углеводов, например пентозофосфатный, исследованный В. А. Энгельгардтом, О. Варбургом и Ф. Дикенсом. Предполагается, что основное значение пентозного цикла - снабжение живого организма пентозами, необходимыми для синтеза нуклеиновых кислот и накопления биомассы.
Основная роль в синтезе белка принадлежит нуклеиновым кислотам, которые образуют протоплазму живой клетки. В состав их входят пуриновые (аденин и гуанин) или пиримидиновые основания (цитозин, урацил, тимин), сахар пентоза и фосфорная кислота. Существует два типа нуклеиновых кислот - рибонуклеиновая (РНК) и дезоксирибонуклеиновая (ДНК), которые различаются между собой по химической природе входящего в их состав сахара и присущим им функциям.
РНК является сложным полимерным соединением, молекулярная масса которого составляет от 18 тыс. до 2 млн. Молекула РНК представляет собой цепь, имеющую спиральное строение. Она состоит из большого количества нуклеотидов. В состав ее входит урацил, а сахарным компонентом является d-рибоза. РНК содержится в цитоплазме и ядре. Строение ее специфично для каждого организма. РНК различается количеством и последовательностью нуклеотидов. РНК играет большую роль в синтезе белка.
По характеру роли, выполняемой в процессе синтеза биомассы, различают три основных вида РНК: рибосомная, транспортная и информационная.
Рибосомная РНК имеет большую молекулярную массу, осуществляет синтез белка в рибосоме.
Транспортная РНК (т-РНК) имеет небольшую молекулярную массу (23 000-28 000); т-РНК переносят аминокислоты в рибосому, где происходит синтез белков. Каждой аминокислоте соответствует своя т-РНК.
Информационная, или матричная, РНК (и-РНК) имеет молекулярную массу 500 000 и более, обладает высокой активностью - быстро синтезируется и распадается. Она осуществляет передачу в рибосому информации, зашифрованной в молекуле ДНК.
ДНК представляет собой соединение, во многом сходное с РНК, но с более высокой молекулярной массой (4-8 млн.). ДНК состоит также из нуклеотидов, в состав которых входят пуриновые и пиримидиновые основания, углевод и фосфорная кислота.
Различие в строении этих кислот состоит в том, что в состав ДНК входит дезоксирибоза и основание тимин, а в состав РНК - сахар рибоза и основание урацил.
По данным Д. Уотсона и Ф. Крика, молекула ДНК представляет собой двойную завитую спираль, как бы две пружины, вставленные одна в другую и соединенные водородными связями. Каждая цепочка представляет собой полинуклеотид. Азотистые основания, входящие в состав двух спиральных цепочек ДНК, соединяются между собой водородными связями в соответствии с правилом Чаргаффа: аденин одной полинуклеотидной цепочки с тимином другой цепочки, а гуанин с цитозином.
Последовательность расположения оснований в одной цепи может быть любой, но в другой цепи основания будут располагаться в последовательности, определяемой расположением оснований в первой цепи, т. е. аденин - гуанин, гуанин - аденин и тимин - цитозин, цитозин - тимин.
В свете последних достижений в биологии клетки синтез белков происходит в мельчайших субклеточных структурах, называемых рибосомами. Рибосомы обнаружены в ядрах, митохондриях, цитоплазме.
Синтез белка осуществляется за счет энергии, выделяющейся при дыхании и брожении. В улавливании и использовании этой энергии клеткой важнейшую роль играют макроэргические соединения, в первую очередь аденозинтрифосфорная кислота (АТФ). Ферментативный синтез белка в живых системах начинается с процесса активирования аминокислот. Для каждой аминокислоты имеется свой специфический фермент, активирующий именно данную кислоту. Процесс синтеза белка в рибосоме схематично изображен на рис. 3 (по А. С. Спирину).
Таким образом, разнообразные экспериментальные данные, накопленные наукой за последние годы, указывают на то, что в живой клетке процесс биосинтеза белка неразрывно связан с процессом биосинтеза нуклеиновых кислот, которые, в свою очередь, синтезируются благодаря каталитическому действию специфических белков-ферментов.
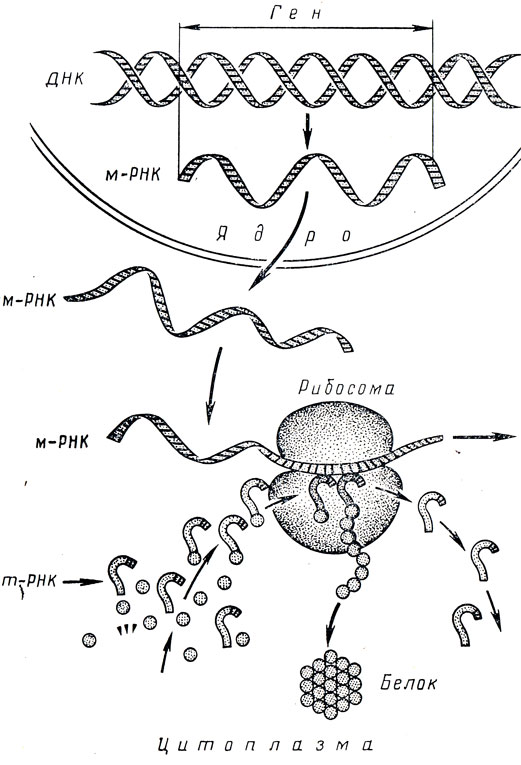
Рис. 3. Биосинтез белка в рибосоме (по А. С. Спирину)
Данные о механизме синтеза белка все время расшифровываются, уточняются и дополняются.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://knigakulinara.ru/ 'Библиотека по кулинарии'